Asal Usul Relung dan Spesies di Dunia Bakteri
Asal Usul Relung dan Spesies di Dunia Bakteri – Relung adalah ruang untuk unit seleksi biologis, dari sel hingga komunitas yang kompleks. Dalam arti luas, “spesies” adalah unit biologis individuasi. Relung tidak ada tanpa organisme individu, dan setiap organisme memiliki relung.
Asal Usul Relung dan Spesies di Dunia Bakteri
hospitalmicrobiome – Kami menggunakan “ceruk” dalam pengertian Hutchinsonian sebagai abstraksi dari ruang lingkungan multidimensi yang dicirikan oleh berbagai kondisi, baik biotik maupun abiotik, yang rentang kuantitatifnya menentukan tingkat pertumbuhan positif atau negatif dari individu mikroba, biasanya suatu spesies, tetapi juga bagian dari komunitas spesies yang terkandung dalam ruang ini.
Organisme mikroba (“spesies“) terus melakukan diversifikasi, dan diversifikasi (radiasi) semacam itu bergantung pada kemungkinan membuka relung yang belum tereksploitasi atau yang kurang tereksploitasi.
Eksploitasi ceruk sering menyiratkan “konstruksi ceruk, ” ketika ceruk yang dijajah berkembang seiring waktu, memunculkan subniche potensial baru, sehingga memengaruhi pemilihan serangkaian varian baru dalam keturunannya. Evolusi relung dan organisme merupakan hasil dari proses interaksi timbal balik yang membentuk satu kesatuan proses.
Baca Juga : Kita Tidak Pernah Sendiri: Hidup dengan Mikrobiota Manusia
Diversifikasi mikroba sentrifugal memperluas batas relung spesies sementara proses sentripetal atau kohesif terjadi secara bersamaan, dimediasi oleh transfer gen horizontal dan peristiwa rekombinatorial, memadatkan semua informasi yang diperoleh selama spesialisasi diversifikasi menjadi “organisme baru” (kemungkinan spesies masa depan), sehingga menciptakan ceruk yang lebih kompleks, di mana keegoisan organisme baru membentuk “kekuatan homeostatik” yang membatasi variasi relung.
Setelah daya dukung penuh ceruk telah tercapai, isolasi reproduktif terjadi, karena tidak ada organisme asing yang dapat mengungguli populasi/komunitas yang ada, sehingga memfasilitasi spesiasi. Dalam kasus individualisasi-spesiasi mikrobiota, kontribusinya terhadap struktur usus hewan adalah sejenis “konstruksi relung”, hasil dari pembicaraan silang antara relung (inang) dan mikroorganisme. Terakhir, ada paralelisme antara hierarki relung dan individu mikroba.
Meningkatnya efek antropogenik pada biosfer (seperti globalisasi) dapat mengurangi keragaman relung dan individu bakteri, dengan potensi munculnya multispesialis yang sangat menular (yang pada akhirnya merusak) yang dihasilkan dari homogenisasi mikrobiosfer, kemungkinan yang harus dieksplorasi dan dicegah. karena tidak ada organisme asing yang dapat mengungguli populasi/komunitas yang ada, sehingga memudahkan spesiasi.
Dalam kasus individualisasi-spesiasi mikrobiota, kontribusinya terhadap struktur usus hewan adalah sejenis “konstruksi relung”, hasil dari pembicaraan silang antara relung (inang) dan mikroorganisme.
Terakhir, ada paralelisme antara hierarki relung dan individu mikroba. Meningkatnya efek antropogenik pada biosfer (seperti globalisasi) dapat mengurangi keragaman relung dan individu bakteri, dengan potensi munculnya multispesialis yang sangat menular (yang pada akhirnya merusak) yang dihasilkan dari homogenisasi mikrobiosfer, kemungkinan yang harus dieksplorasi dan dicegah. karena tidak ada organisme asing yang dapat mengungguli populasi/komunitas yang ada, sehingga memudahkan spesiasi.
Dalam kasus individualisasi-spesiasi mikrobiota, kontribusinya terhadap struktur usus hewan adalah sejenis “konstruksi relung”, hasil dari pembicaraan silang antara relung (inang) dan mikroorganisme. Terakhir, ada paralelisme antara hierarki relung dan individu mikroba. Meningkatnya efek antropogenik pada biosfer (seperti globalisasi) dapat mengurangi keragaman relung dan individu bakteri, dengan potensi munculnya multispesialis yang sangat menular (yang pada akhirnya merusak) yang dihasilkan dari homogenisasi mikrobiosfer, kemungkinan yang harus dieksplorasi dan dicegah. Dalam kasus individualisasi-spesiasi mikrobiota, kontribusinya terhadap struktur usus hewan adalah sejenis “konstruksi relung”, hasil dari pembicaraan silang antara relung (inang) dan mikroorganisme. Terakhir, ada paralelisme antara hierarki relung dan individu mikroba. Meningkatnya efek antropogenik pada biosfer (seperti globalisasi) dapat mengurangi keragaman relung dan individu bakteri, dengan potensi munculnya multispesialis yang sangat menular (yang pada akhirnya merusak) yang dihasilkan dari homogenisasi mikrobiosfer, kemungkinan yang harus dieksplorasi dan dicegah. Dalam kasus individualisasi-spesiasi mikrobiota, kontribusinya terhadap struktur usus hewan adalah sejenis “konstruksi relung”, hasil dari pembicaraan silang antara relung (inang) dan mikroorganisme. Terakhir, ada paralelisme antara hierarki relung dan individu mikroba.
Meningkatnya efek antropogenik pada biosfer (seperti globalisasi) dapat mengurangi keragaman relung dan individu bakteri, dengan potensi munculnya multispesialis yang sangat menular (yang pada akhirnya merusak) yang dihasilkan dari homogenisasi mikrobiosfer, kemungkinan yang harus dieksplorasi dan dicegah. ” hasil crosstalk antara niche (inang) dan mikroorganisme. Terakhir, ada paralelisme antara hierarki relung dan individu mikroba. Meningkatnya efek antropogenik pada biosfer (seperti globalisasi) dapat mengurangi keragaman relung dan individu bakteri, dengan potensi munculnya multispesialis yang sangat menular (yang pada akhirnya merusak) yang dihasilkan dari homogenisasi mikrobiosfer, kemungkinan yang harus dieksplorasi dan dicegah. ” hasil crosstalk antara niche (inang) dan mikroorganisme. Terakhir, ada paralelisme antara hierarki relung dan individu mikroba. Meningkatnya efek antropogenik pada biosfer (seperti globalisasi) dapat mengurangi keragaman relung dan individu bakteri, dengan potensi munculnya multispesialis yang sangat menular (yang pada akhirnya merusak) yang dihasilkan dari homogenisasi mikrobiosfer, kemungkinan yang harus dieksplorasi dan dicegah.
Relung adalah ruang untuk unit seleksi biologis. Tinjauan berpendirian ini bermaksud untuk membahas asal usul, pengembangan, dan evolusi relung dari sudut pandang ekologi, sebagai alat rahim yang mengarah pada pengiriman individu mikroba (unit seleksi), dari spesies ke komunitas yang stabil di dunia bakteri. Keberadaan rahim memiliki sedikit arti tanpa organisme berkembang di dalamnya. Demikian pula, relung tidak ada artinya tanpa organisme yang berkembang di dalamnya, mengingat konsep relung di dunia pra-biotik tidak ada artinya. Ini adalah organisme individu yang hidup yang memberikan identitas ceruk. Misalnya, berbagai pulau berbatu di Galapagos bukanlah “relung” untuk berbagai spesies burung kutilang sebelumnya .kedatangan stokastik burung kutilang (tidak berdiferensiasi) dari benua jutaan tahun yang lalu. Mereka menjadi “relung” karena spesiasi burung finch. Begitu ada korespondensi antara spesies dan ceruk, relung itu bertahan sedemikian rupa sehingga bahkan jika semua anggota spesies punah secara lokal, relung kosong akan mengenali atau dikenali oleh gelombang masuk baru individu-individu dari relung spesifik yang sama. spesies, tertarik oleh area aksesibilitas historis ini.
Sebenarnya, asal kata ”niche” berkaitan dengan bahasa Latin ” nidus” sarang, menyediakan kondisi yang mendukung perkembangan suatu organisme. Burung membangun sarangnya, yang penting untuk perkembangan spesies. Akhirnya, spesies burung lain (mungkin terkait) mungkin mengenali sarang alien yang cocok untuk perkembangannya sendiri (burung perampok sarang). Perhatikan bahwa konstruksi sarang (relung) seringkali membutuhkan cabang-cabang pohon tertentu di lingkungan tertentu (biasanya “bersejarah”, yaitu, yang telah digunakan oleh generasi sebelumnya) yang dapat menyediakan makanan bagi anak-anaknya, bertindak sebagai “sarang”. ” untuk sarang, sehingga menciptakan citra intuitif hierarki sarang. Bagaimana hal ini berlaku untuk spesiasi bakteri? Mengutip Jacques Monod dari perspektif anti-reduksionistik, “apa yang benar untuk burung juga berlaku untuk bakteri.”
Kami tentu dapat memperluas pertimbangan ini, mengingat struktur fraktal alam, mengingat bahwa “spesies” adalah kesatuan biologis individuasi (membangun, mengembangkan individu) dan seleksi, dan ceruk (sarang) adalah unit fisikokimia spasial yang sesuai dari individuasi dan seleksi. Padahal, sebelum abad ke-16, arti istilah “spesies” adalah “kelas (sesuatu) yang berbeda berdasarkan karakteristik umum,” yaitu kategori sesuatu yang dapat dihargai (specere , “untuk melihat”) sebagai entitas individu. Istilah spesies (biologis) modern diciptakan oleh John Ray pada abad ke-17 dan diterapkan secara luas oleh Carl Linnaeus pada abad ke-18, mengacu pada tumbuhan dan hewan. Di zaman kita, kebangkitan tertentu dari makna lama diperlukan, memungkinkan untuk mencakup entitas individu biologis di bawah dan di luar definisi klasik spesies.
Konsep spesies bakteri sangat sulit dipahami. Seperti halnya burung perampok sarang, beberapa jenis organisme bakteri kerabat cocok di ceruk yang sama. Oleh karena itu telah diusulkan bahwa “relung” menggambarkan batas unit ekologi mikroba, tidak harus spesies tetapi organisme spesifik relung yang setara secara ekologis, “ekotipe,” sehingga spesies yang dinamai, disusun oleh ekotipe, dapat menyerupai apa yang kita sekarang dipahami sebagai genus ( Cohan, 2002 ; Ward et al., 2006 ). Memang, komponen spesies bakteri terkait secara filogenetik dan berbagi eko-fenotipe yang serupa ( Rossello-Mora dan Amann, 2001).). Namun, yang dikaitkan dalam ulasan ini dengan “spesies” dapat sama-sama dikaitkan dengan individu evolusioner lainnya di tingkat yang lebih tinggi (genus atau bahkan komunitas terintegrasi dari spesies yang berbeda) dan tingkat hierarki yang lebih rendah (subspesies, klon) sesuai dengan konsep spesies yang berbeda ( Palmer dkk., 2019). Minat baru dalam “mikrobiologi ceruk” dipupuk oleh meningkatnya kebutuhan untuk menentukan risiko yang terkait dengan peningkatan efek antropogenik pada mikrobiosfer. Dalam perspektif One Health dan kesehatan Global, kesehatan manusia, dan kesehatan sebagian besar kehidupan terestrial, bergantung pada keseimbangan kompleks yang didominasi oleh konglomerat, efek integratif mikrobiosfer. Efek tersebut mencakup dari mempertahankan kehidupan produsen utama hingga penyebaran resistensi terhadap agen antimikroba ( Baquero et al., 2019 ).
Konsep Niche Bakteri
Istilah “niche” diciptakan pada tahun 1913 oleh Grinnell dan Swarth ketika membahas spesiasi burung finch Galapagos ( Grinell dan Swarth, 1913). dan dikandung sebagai rumah bagi satu spesies atau akhirnya subspesies. Kita harus menyadari bahwa istilah “ceruk” mencerminkan abstraksi, dalam arti metafora, dan akibatnya upaya untuk definisi yang tepat tergantung pada sudut pandang dan niat ilmuwan, yang mengakibatkan kurangnya kejelasan secara keseluruhan ketika diterapkan. untuk tujuan yang berbeda. Di zaman kita, niche secara bersamaan adalah ruang (tetapi ruang cair, bukan gelembung), pemandangan di mana kondisi abiotik, non-interaktif, dan akhirnya variabel lingkungan tertentu terjadi, area di mana fungsi yang berasal dari biotik, termasuk interaksi biologis pada hierarki yang berbeda. sisik terus-menerus mengubah kondisi lokal (seperti mengkonsumsi atau memproduksi nutrisi), dan lokasi yang fleksibel di mana beberapa organisme menetap (bertahan hidup, bereproduksi) dan organisme lain bermigrasi ke daerah yang dapat diakses.
Relung Grinellian mengacu pada ruang multidimensi yang dibentuk oleh ansambel variabel abiotik independen, tidak atau lambat interaktif dan tidak dapat dikonsumsi yang mempengaruhi organisme (variabel scenopoetic, yang berarti variabel yang tidak berinteraksi). Relung Eltonian mengacu pada ansambel lokal interaksi biotik cairan pada skala seluler, termasuk variabel bionomik yang relatif statis sebagai kompetisi untuk nutrisi atau mutualisme (Soberon, 2007 ; Soberon dan Nakamura, 2009 ). Perhatikan bahwa semua pandangan yang berbeda ini sebagian besar lahir untuk mendekati masalah biogeografis, termasuk identifikasi area distribusi (seringkali mengenai tumbuhan dan hewan), tetapi pada kenyataannya mereka tidak dapat dianggap saling eksklusif. Bagaimana gagasan heuristik ini dapat diterapkan pada relung mikroba, yang tersebar luas, tetapi juga berbutir halus dan terputus-putus?
Untuk tujuan tinjauan ini, berfokus pada relung mikroba, makna relung sebagian besar didasarkan pada yang diusulkan oleh George Evelyn Hutchinson (1903–1991) ( Hutchinson, 1957 ; Holt, 2009) sebagai abstraksi dari ruang lingkungan multidimensi yang dapat dicirikan oleh berbagai kondisi, baik biotik (sebagai interaksi bakteri) dan abiotik (biotope), yang rentang kuantitatifnya menentukan laju pertumbuhan positif atau negatif dari unit evolusi (biasanya a spesies tetapi juga komunitas spesies) yang terdapat dalam ruang tersebut. Konsep relung dan biotope Hutchinsonian tetap sangat berguna dalam biogeografi, ekologi, epidemiologi, dan evolusi tetapi dapat digunakan untuk mensimulasikan efek pada spesies dan komunitas mikroba selama pembentukan relung dan evolusi di bawah prediksi perubahan lingkungan lokal dan global ( Colwell dan Rangel, 2009 ) ).
Kondisi ceruk Hutchinsonian mengingatkan “fenotipe” organisme, dan ciri-ciri “fenotipe ceruk” harus sesuai (seperti dalam “metafor cermin”) dengan organisme yang mampu bertahan atau tumbuh di dalam ceruk. Lebih dari 25 tahun yang lalu, salah satu penulis ulasan ini (FB) menyebut ansambel yang tidak dapat dipisahkan dari tautan ini “ecobiome” ( Baquero, 1994 ) memperluas definisi pertama “ecobiome” sebagai ansambel kondisi biotik dan abiotik dari ceruk ekologis ( Polunin, 1984 ). Konsep esensial adalah “permukaan respons niche” yang diturunkan dari fungsi biologi populasi r(e) , yang didefinisikan sebagai ketergantungan laju pertumbuhan populasi ( r) pada ansambel komponen biotik dan abiotik lingkungan ( e ) ( Holt, 2020 ). Oleh karena itu, ceruk secara bersamaan merupakan lingkungan dan membran individu biologis, yang merupakan dualitas ontologis yang tidak dapat dipisahkan. Relung “ditemukan” (dibuka) oleh varian mikroba. Lingkungan tertentu di mana pemilihan varian tertentu berlangsung diubah menjadi “ceruk”, yang kemudian mengindividualisasi (spesiasi, dalam arti luas) organisme.
Relung: Struktur dan Variasi
Relung memiliki komposisi internal yang bertanggung jawab atas “respon relung” ( Maguire, 1973 ), mengingat bahwa ia terdiri dari ansambel multidimensi dari kondisi lokal yang terdistribusi secara heterogen di dalam relung, sering kali membentuk gradien. Dalam hipotesis sentralitas ceruk, harus ada pusat geometris teoretis (“pusat” atau “inti”), yang mewakili kualitas keseluruhan rata-rata ceruk. Di dalam relung, harus ada tempat di mana kombinasi kondisi spesifik tertentu optimal untuk menyediakan pengait ke inokulum langka (propagulum) dari mikroorganisme tertentu, sehingga dapat bertahan dan tumbuh, idealnya di atau di dekat inti relung. . Secara hipotetis, kelimpahan spesies yang beradaptasi dengan niche di situs ini harus maksimal ( Yañez et al., 2020). Situasi ini mengingatkan lanskap kebugaran Sewall Wright, dengan puncak optimal di mana genotipe berlimpah ( Wright, 1932 ). Seperti di lanskap Wright, inti relung tidak perlu teratur (lingkaran) dan dapat berbentuk bulat telur atau bahkan asimetris ( Holt, 2020 ), tanpa puncak tunggal melainkan punggungan tinggi, mengingat satu atau lebih kondisi di ceruk bisa lebih stabil atau dominan daripada yang lain dalam memberikan kelimpahan semaksimal mungkin ( Holt, 2020 ; Osorio-Olvera et al., 2020 ).
Namun, kondisi yang mengonfigurasi inti relung mungkin tidak stabil, yang membuatnya sulit untuk mengidentifikasi area kelimpahan maksimum organisme tertentu. Oleh karena itu, inti mengikuti lintasan daripada yang dapat direpresentasikan lebih sebagai lokus geometris daripada sebagai sentrum. Populasi mikroba diharapkan untuk melakukan perjalanan terus-menerus untuk mencari ceruk inti fundamental mereka (atau yang lebih dekat), dan variasi bersama ini, yang dimediasi oleh peristiwa penularan, mengurangi pemutusan hubungan antara spesies dan lingkungan. Dalam lingkungan yang tidak stabil, kelimpahan harus diukur sebagai kepadatan rata-rata di sepanjang lokus geometris dan waktu. Relung dibatasi oleh “membran” eksklusif yang memfasilitasi isolasi reproduksi organisme.
Pergerakan (tidak jarang osilasi) dari pusat geometris dari ceruk asli di bawah berbagai kondisi mengubah topologi parametrik relung, yang menghasilkan pembukaan ruang dengan kelimpahan rendah (relung periferal) hingga varian mikroba akhirnya mampu menangani atau mengeksploitasi suboptimal ini. kondisi, mengubahnya menjadi neo-inti untuk varian genotipe dan memfasilitasi keabadian dan pertumbuhannya. Meskipun situasi ini seharusnya menciptakan persaingan klonal dengan organisme leluhur pionir, variabilitas ceruk memastikan koeksistensi dan meningkatkan kemungkinan spesies (sebagai ansambel varian mikroba) untuk mencapai daya dukung penuh relung.
Relung dengan tingkat kompleksitas dan stabilitas yang berbeda jelas mempengaruhi evolusi populasi bakteri. Sebuah “ceruk datar” dengan sedikit komponen, dominasi kuat dari satu atau beberapa kondisi, sedikit dan gradien lembut ( Colwell dan Rangel, 2009) dan kondisi stabil di seluruh ceruk harus memiliki efek yang berbeda pada evolusi spesies mikroba daripada relung lainnya. Misalnya, jika suhu (atau osmolaritas) sangat tinggi, ceruk cenderung datar, karena komponen ini menciptakan gaya selektif tunggal (atau dominan). Jika ceruknya datar dan besar, pemilihan varian mikroba (dan diversifikasi dan spesiasi yang dihasilkan) harus berkurang. Sebaliknya, relung dengan kelimpahan dan kompleksitas tinggi tetapi homeostasis yang sangat efisien (misalnya, sitoplasma sel eukariotik) memastikan “kerataan” yang serupa, mengingat homeostasis mengoordinasikan/menyederhanakan keragaman komponen, menghasilkan hampir satu komponen dengan efisiensi selektif .
Stabilitas Niche dan Radiasi Adaptif
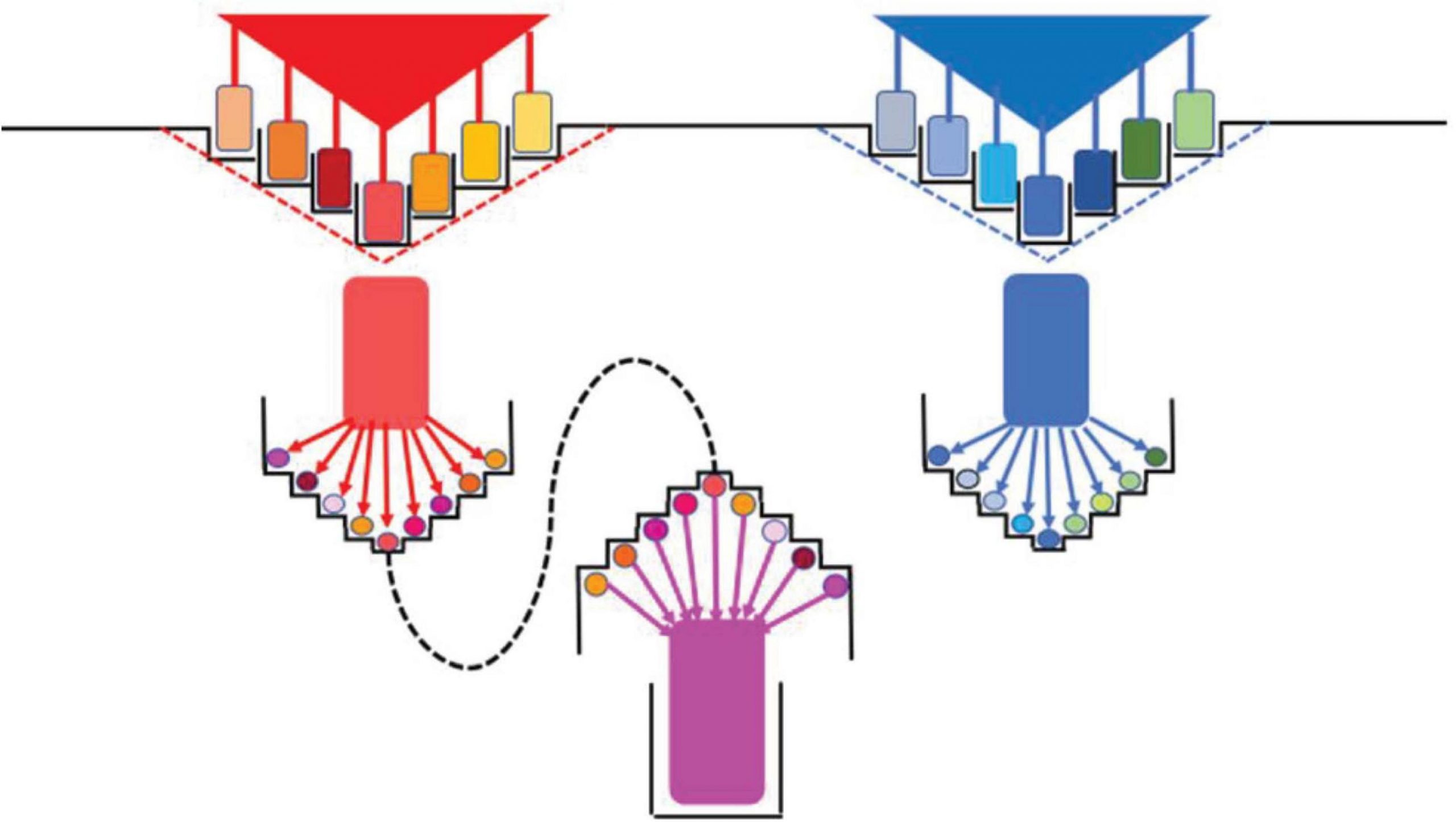
Cukup “stabilitas rata-rata” dari kesetiaan inti ceruk-organisme sangat penting untuk proses spesiasi. Jika variasi niche mendukung diversifikasi spesies sebagai ansambel varian subspesifik, proses spesiasi membutuhkan “kompartemen” (sarang stabil) yang memfasilitasi isolasi reproduksi organisme. Stabilitas niche juga memastikan replikasi yang efektif dan kemungkinan menjelajahi lingkungan tetangga yang stabil untuk diubah menjadi niche. Proses eksplorasi, penemuan, dan kolonisasi neo-relung oleh varian fenotipik yang menghasilkan garis keturunan yang berkembang pesat ini disebut “radiasi adaptif” ( Gavrilets dan Vose, 2005 ; Gillespie dan Parent, 2014 ).). Paul Rainey dkk. melakukan eksperimen mani yang menunjukkan munculnya garis keturunan baru (spesies potensial) di lingkungan yang stabil ( Rainey dan Travisano, 1998 ). Dalam labu berisi kultur cair yang dikocok, inokulum dari jenis bakteri tampaknya tetap homogen secara genetik dan ekologis. Namun, jika inokulum disemai ke dalam labu tanpa pengocokan, kompartementalisasi kondisi kultur terjadi secara spontan, menciptakan sub-lingkungan yang terstruktur secara spasial [misalnya, kandungan oksigen rendah di bagian bawah, berlimpah di permukaan, dengan kultur udara tertentu. interaksi permukaan kaca ( Jerdan et al., 2019)]. Dalam waktu singkat (hari), “genotipe spesialis” (dalam pelat kultur) muncul dan secara istimewa menempati berbagai kompartemen ketika diinokulasi dalam labu baru. Genotipe mudah dikenali oleh kekhasan koloni yang mereka bentuk ketika tumbuh di piring agar. Beberapa mutasi telah ditemukan bertanggung jawab atas proses radiasi adaptif ini, beberapa lebih efektif daripada yang lain ( McDonald et al., 2009 ). Radiasi adaptif juga telah diamati dalam model lingkungan konstan ( Maharjan et al., 2006 ), tetapi sulit untuk mengesampingkan keberadaan “kompartemen mikro” di lingkungan stabil yang tampaknya tidak terstruktur, terutama jika perangkat kompleks (seperti chemostat) digunakan .
Kapasitas divergensi bakteri tanpa memerlukan spesialisasi ekologi di beberapa relung dipelajari dengan kultur kemostat Escherichia coli dengan sumber nutrisi terbatas ( Maharjan et al., 2006 ). Populasi klon awalnya menyebar ke beberapa kelompok dengan peningkatan penggunaan nutrisi, bersaing tetapi berbagi ceruk yang sama (tidak ada seleksi berkala), dan dengan demikian dianggap milik “ekotipe tunggal.” Eksplorasi multi arah ruang kebugaran merupakan faktor yang diremehkan dalam diversifikasi bakteri. Studi in vitro ini memiliki korespondensi dengan kehidupan nyata, dan spesiasi simpatrik dalam genus Vibrio laut didasarkan pada diversifikasi awal dalam mikroniche air (Shapiro dkk., 2012 ; Friedman et al., 2013 ).